Krysy přicházejí postranní paleta barev a vzorů, od pevné látky černé k white, od tmavý zhnědne ohřívat opaluje se k krémům, od břidlice šedé k bledě blue. Krysy mohou zobrazovat záhony white ten rozsah od malého hrudního bodu k břichu blotch k bílé s zabarvenou hlavou a dál. Variace je ohromná! Kde tito natírají barvy a vzory přijít z? Jak být různá barviva dělala a začleňovala do rostoucí kožešiny? Jak dělat mutace podél barvivové výrobní stezky produkují různé barvy, které my vidíme v krysách?
- Barvivový základ
- Mutace v barvivové buňce distribuce
- Bílá místa a body: zahalený, všímá si smrtelného a bílého nanášení mutace
- Mutace v melanosome formace
- Mutace v barvivu doprava a nános
- Mutace v barvivu syntéza
- Albín a siamese: mutace v chinchilla gen
- Pink-eyed roztok
- Brown
Popření: Já jsem biolog, ne krysí chovatel a můj zájem na krysích barvách pochází z mé fascinace biologií zabarvení. Proto, toto je ne článek o krysích standardech! Jestliže vy hledáte krysí barevné standardy, popisy, a jejich spojil Mendelian zápisy, podívat se na jednoho z mnoho websajtů přepychových krysích organizací takových jak AFRMA, RMFE, NFRS, a AusRFS k jménu správnému několik.
Ne všechny krysí barvy jsou zahrnuté tady, protože já jsem soustředil mou pozornost na mutacích jehož buněčné důsledky jsou známé. Tam být mnoho jiné barvy jehož konkrétní příčiny nejsou přesto známé.
Slovo o barevných jménech ... Já jsem občas zahrnoval barevná jména pro účely ilustrace. Poznámka, nicméně, že taková barevná jména často mění se mezi různými zeměmi a krysími organizacemi. Stejná barva může být nazvaná různými jmény a zvláštním jménem může odkazovat se na různé barvy, v závislosti na kom vy mluvíte k. Také, zvláštní barva může mít množství možných základních příčin tak dbát na opatrnost když vyvozuje z tohoto dokumentu k vašim oblíbeným krysám. U a velký, já jsem snažil se k volu jasnému krysy standardy a " úředník " barví jména a má místo toho soustředěný na mé zájmové oblasti: biologie zabarvení.
Barvivový Základ
Barviva jsou vyrobená v buňkách nazvaných melanocytes. Během vývoje embrya, buňky, které stanou se melanocytes (tito předchůdcové jsou nazvaní melanoblasts) stěhovat se od hřbetní oblasti -- od aktivní oblasti odprejskne zpět embrya nazvaného nervový hřeben -- k zbytku těla. Tito předchůdcové začnou s pozicemi u základu vlasových folikulů. Oni také stanou se integrálními částmi jiných orgánů takových jako oko, vnitřní ucho a jiné nervové systémové (Číslo 1 ). Jestliže stěhování těchto buňek je přerušené, pak některé oblasti těla nebudou mít melanocytes a tyto oblasti těla budou produkovat bílé vlasy.
| Číslo 1. Během vývoje embrya, melanocyte předchůdcové stěhují se od nervový hřeben. Tyto nervové hřebenové buňky rozlišují do mnoho různá buňka píše, zahrnující melanocytes. Nepořádky v melanocyte stěhování a rozdílnost zahrnovat zahalený, nanášející smrtelný, a bílé nanášení. |
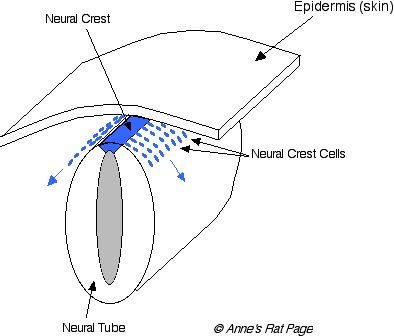
Melanoblasts rozlišují do melanocytes, který žít v kůži a u základu každých vlasů a produkčního melaninu barviva. Menanin barvivové částečky, volaný granules, být včleněn do pěstování vlasového (Čísla 2 ).
| Číslo 2. Typický Vzorek vlasů a váčku. Melanocytes syntetizuje melanin granules, které jsou včleněny do pěstování vlasového (kliku na obrazu pro větší obraz ). |
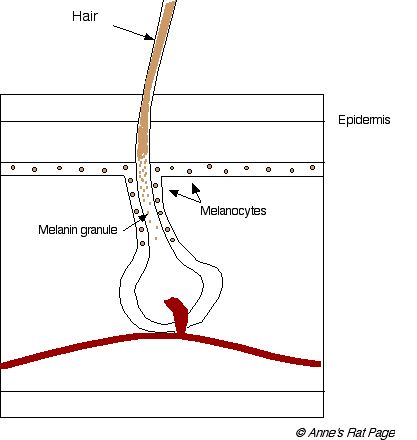
V melanocyte ...
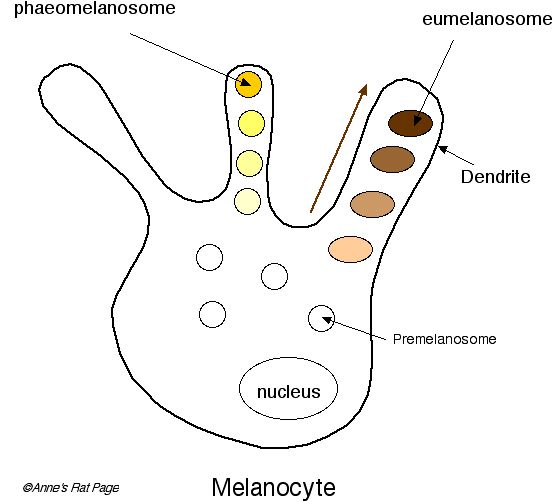
Barvivové částečky, volaný granules, být vysyntetizovaný v malém vesicles v melanocytes. Tyto vesicles jsou nazvané melanosomes. Dva druhy barviva jsou vytvořené v melanosomes: žlutá červená barviva nazvaná phaeomelanins a hnědá černá eumelanins.
Tam jsou vlastně oddělené melanosomes pro dva druhy barviva. Phaeomelanosomes specializovat se v dělání žlutého červeného phaeomelanins. Eumelanosomes specializovat se v dělání hnědého černého eumelanins. Když melanocyte přepne mezi produkováním bledých a tmavých barviv v jediných vlasech, účinek je svazoval vlasy jak viděl v agouti allele.
Melanosomes produkuje barviva v melanocyte. Melanosomes začne jejich cestu v středu melanocyte buňky, a stěhovat se k vnějšímu okraji buňky, přes projekce nazvané dendrites (Číslo 3 ).
[Pamatovat si, melanocytes jsou vlastně druh nervové buňky, přijde z nervového hřebenu, tak oni mají některé neuron-like vlastnosti takový jak dendrites ].
U vnějšího okraje buňky, melanosomes uvolní jejich barvivo, který pak incoporated do obklopování keratinocytes a komína rostoucích vlasů.
| Číslo 3. Melanocytes obsahuje melanosomes: phaeomelanosomes a eumelanosomes, který syntetizovat bledá a tmavá barviva. Přepíná Mezi bledým a tmavým barvivem výroba na jediných vlasech produkuje odstupňované vlasy, nazvaný agouti. Melanosomes stěhuje se k základu dendrites. Odtamtud oni jsou transportovaní k dendrite tipům. Nepořádky v melanosome formace v krysách zahrnovat red-eyed prohlížela si roztok. |
Jak přece melanosomes stěhují se od středu k okraji melanocyte? Melanosomes je vlastně spojený s kostrou z microtubules, a být přenesený zvýšit dendrites na žebříkách actin. Malé " nohy " , které jdou zvýší actin vlákna jsou nazvaná myosin 5 (Číslo 4 ).
| Číslo 4. Melanosomes je spojený s odvětvováním microtubule stromu v melanocyte. Postoupit do dendrite, oni vážou se k actin vláknům s myosin 5, a myosin 5 " jde " je do buněčného okrajového (kliku na obrazu pro větší obraz ). Nepořádky v melanosome doprava zahrnovat roztok allele. |
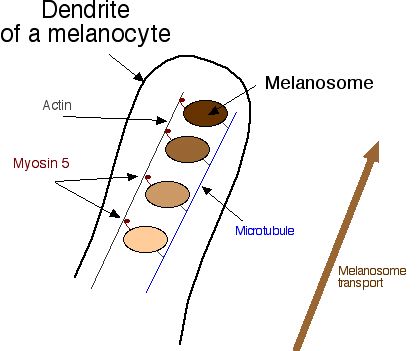
V melanosome ...
V melanosomes, savci dělají dva druhy barviv: eumelanins, který rozsah od tmavě brown odmítnout, a phaeomelanins který rozsah od červené k yellow.
Oba druhy barviva jsou vyrobené z stejného začínajícího produktu, amino kyselina tyrosine, a nejprve několik kroky jejich výroby jsou stejný.
Enzyme tyrosinase (kódovaný genem nazvaným chinchilla) přestaví tyrosine, který je bezbarvý, do dopaquinone, také bezbarvý. Phaeomelanins je vyrobený z těchto.
Dělat eumelanin, dopaquinone je přeměněn na dopachrome. Dopachrome může vzít dvě cesty k eumelanin. V první stezce, dopachrome je přeměnil na 5, 6 dihydroxyindole, který je hnědý, a pak do komplexu, který quinone separace enzyme tyrosinase příbuzná bílkovina já (TYRP1, produkoval u brown gen ). V druhé stezce to je přeměněno na DHCA u dopachrome tauromerase (DCT) a odtamtud k komplexu quinones prostřednictvím TYRP1 znovu. Komplexní quinones pak polymerized do eumelanin, který je černé (Číslo 5 ).
| Číslo 5. Zjednodušená syntéza melanins phaeomelanin a eumelanin. Nepořádky v barvivová syntéza zahrnovat chinchilla (kódy tyrosinase) a brown (kódy TYRP1 ). (klik na obrazu pro větší obraz ). Kompromis mezi syntézou phaeomelanin a eumelanin podél komína rostoucích vlasů je zprostředkovaný u agouti allele. |
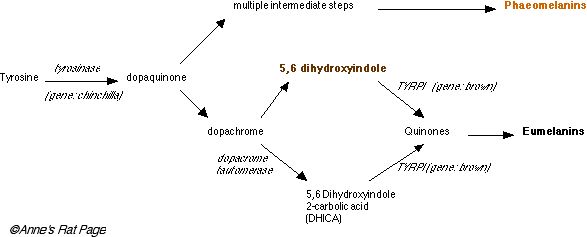
Phaemelanosomes a eumelanosomes jsou vlastně docela různý vnitřek. Phaeomelanosomes je kulatý ve tvaru a je docela primitivní, postrádání TRP1, TRP2, a p-protein (viděný v pink-eyed prohlížela si roztok). Oni mají jen jednu třetinu stupeň tyrosinase jako eumelanosomes.
Eumelanosomes je důmyslnější, ovál zformovaný melanosomes, který mít TRP1, TRP2, p-protein a tři časy tyrosinase jako phaeomelanosomes.
Závěr
Variace v vlasové barvě mohou vzniknout z rozdílů v syntéze barviva, končí black, hnědé, žluté nebo bezbarvé barvivo granules, nebo v depositu barviva v vlasové šachtě. Rozdíly v barvivovém depositu v vlasech produkují variaci v intenzitě a odstínu barvy. Barva a množství granules může měnit se podél délky jediných vlasů nebo mezitím různých vlasů. Variace v zabarvení přijde z vzorů melanocyte stěhování a finální distribuce. Oblasti s melanocytes stanou se zabarvené, ale oblasti bez melanocytes produkce depigmented, bílé vlasy.
V příštích sekcích já budu zkoumat mnoho různé mutace occuring podél této stezky, a jak tyto mutace dají svah konečné vrchní vrstvě barva. Já budu také zkoumat jiné " vedlejší " účinky těchto mutací a možných protějšků najitých v lidském zabarvení.
Pro velké shrnutí barvivové výroby, vidět pronásledovat kabátový barevný odkaz.
Mutace v Melanocyte Distribuci
Zpoždění v melanocyte stěhování: zahalený alleles, nanášející smrtelný, a bílé nanášení alleles
Úvod
V embryu, záhyb vyvíjí se dole zpět volal nervová trubka, který obsahuje aktivní oblast nazvanou nervový hřeben. Tato oblast dodává barvivové buňky (melanocytes) to stěhovat se všude po tělu.
Specificky, barvivové buňky stěhují se k párům specifických míst na jedné straně těla také jako backline. Tam jsou tři taková místa na hlavě (blízko oka, blížit se k uchu a blížit se k vrcholu hlavy ), a šest míst podél každé strany těla, a několik podél ocasu. Několik barvivové buňky stěhují se k každému těchto míst, kde oni množí se a stěhují se ven, připojí se tvořit větší místa, rozšiřující se srážet dolů nohy a srážet dolů hlavu do okamžiku než oni setkají se pod bradou, a srážet dolů tělo do okamžiku než oni setkají se na břichu (Cattanach 1999 ).
Jakmile barvivové buňky skončily stěhovat se oni zaujmou postoje u základu vlasových folikulů. Tam oni syntetizují barvivo melaninu a vkládají to do rostoucích vlasů. Normálně, všechny váčky mají barvivové buňky spojené s nimi a celá zvířecí kožešina je zabarvená. Ale jestliže žádné barvivové buňky jsou spojovány s váčkem, tam je žádné barvivo v tom že vlasy. Mutace, které ovlivní barvivovou buněčnou distribuci během vývoje embrya určují kterého části těla mají barvivové buňky, a od této doby produkovat barvivo, a které části mají žádné barvivové buňky a produkují depigmented vlasy.
Barvivové buňky také stěhují se k duhovce a sítnici oka. Jestliže duhovka nemá barvivové buňky, to vypadá červené (v krysách a myších) nebo modří (psi a kočky ). Odd-eyed krysy jsou způsobené stěhováním barvivových buňek k jednomu oku ale ne jiný. (Poznámka: štuková omítka a červené oči z albíni nejsou způsobení nedostatkem barvivového buněčného stěhování, ale neschopností barvivových buňek k produkčnímu barvivu ).
Barvivové buňky stěhují se k vnitřnímu uchu příliš (kochlea a stria vascularis ), kde oni hrajíundefined ale zásadní role v udržujícím sluchu. Jestliže vnitřní ucho nemá barvivové buňky, jednotlivec může být hluchý.
Barvivové buňky také stěhují se k mozku, k oblastem takovým jako substantia, který nigra (část midbrain, který reguluje náladu, produkuje dopamin a ovládá dobrovolný pohyb ), místo ceruleus (část mozku, který zabývá se stresovou odezvou) také jako jiné oblasti takové jako leptomeninges (blány, které obklopují mozek ), hřbetní kořen ganglia a lebeční ganglia. Nedostatek barvivových buňek k dosahu tyto oblasti mohou mít širokou rozmanitost účinků, takový jak pohybový nepořádek (e.g. záchvaty ), a různorodé účinky na chování a jednotlivce je odezva na stres.
Barvivové buňky jsou proto zapletené do oblastí mozku příbuzného náladě a stresové odezvě. Toto spojení mezi depigmentation a chováním pravděpodobně hrálo roli v zvířecí domestikaci: tím, že vybere pro tameness, chovatelé měnili barvivové buněčné stěhování v vyvíjejícím se nervovém systému, vede k klidnějším zvířatům. Vedlejší účinek tohoto výběru pro chování byl změna v barvivovém buněčném stěhování v kůži, vede k strakatému kabátu. Piebaldness a sdružená poslušnost být najitý v mnoho různí domáčtí druhoví (koně, krávy, psi, kočky, ptáci ). Ve Skutečnosti, výběr divokých zvířat pro krotké chování vede k depigmented oblasti na kožešině jak ukázal se v liškách (Belyaev 1979, Trut 1999) a krysy (Trut et al. 1997 ). Poznámka, nicméně, to jestliže depigmentation je extrém, zvíře může mít neurologická poškození (Grandin 1998 ). Tady je více na kabátová barva, temperament a domestikace.
Barvivové buňky nejsou jediné buňky, které stěhují se od nervového hřebenu. Nervové buňky, které enervate střeva dělají stěhování příliš. Jestliže nutné nervové buňky nedosáhnou konce střeva, zvířecí střeva nemohou fuction vhodně, vyjadřování zvířete neschopného projít kolem feces, který skončí megakolon.
Tam je mnoho mutací to může ovlivnit stěhování buňek od nervového hřebenu. Každá mutace je různá, každý může být upravený jinými geny. Každá mutace může mít strukturu účinků (pleiotropic efekty) na kabátové barvě, chování a smyslová funkce. Toto je proč bílé zbarvení, prohlížet si barevné anomálie, hluchota a megakolon je často najit spolu. Oni jsou celý výsledek zpoždění v buněčném stěhování od nervového hřebenu.
The zahalený allele
The zahalený allele v kryse zdrží stěhování melanocytes od nervového hřebenového (Čísla 1 ). Následně, oblasti nejdalečejší od hřbetního midline -- noh, hruď, břicho -- nemají melanocytes a ty oblasti produkce depigmented, bílé vlasy.
| Číslo 1. Normální a zahalené krysí zárodky, představení dny na kterém melanoblasts dosáhnou pokožky. Zahalené krysy zažijí zpoždění v barvivovém buněčném stěhování (Searle 1968, od Wendt-Wagener 1961) Klik na obrazu dostat větší pohled. |
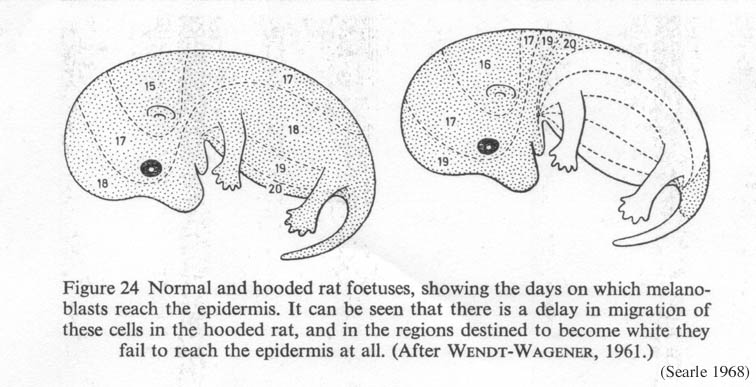
- H = normální, úplně zabarvený kabát
- h = strakatý (zahalený)
Tam být mnoho různé zahalené alleles, které způsobí různé míry zpoždění v melanocyte stěhování: e.g. h (re) (omezený, homozygotní smrtelný ), h (i) (Irský ), h (n) (vrub, nebo opatřený ), h (e) (extrém ), která produkční proměnná množství depigmentation (Robinson 1989 ). Jiné geny ovlivňují přesný nákres depigmented oblastí (Curtis & Dunning 1951 ), takový jak krycí modifikátorové místo h (l) pro dlouho hřbetní pruh a h (s) pro nějaký krátký.
Příklady: Krysy s HH mít normální barvivovou distribuci, která pokrývá jejich celá těla, krysy s Hh částečně depigmented (bílé břicho a zabarvené strany ), známý jak berkshire, a hh krysy jsou zahalený (zabarvená hlava a hřbetní stipe, bílé tělo ).
Lidské analogie: Nejmírnější analogový zahaleného genu je strakatý gen u lidí. Strakatí lidé mají bílý pramen vlasů, žádné barvivo na středu čela a jiných záhonů depigmentation jinde na jejich tělech.
Nanášející smrtelný a bílé nanášení geny
Nervové hřebenové buňky od dalekého konce nervového hřebenu (nazvaného eneterické buňky) stěhovat se k střevu a být zodpovědný za ennervating dvojtečka. Normální melanocyte a eneterický nervový hřeben buněčné stěhování závisí na přítomnosti endolethin B a endolethin B receptory, který regulovat rozdílnost, rozšíření a stěhování melanocytes a eneterických nervových hřebenových buňek během vývoje.
Mnoho mutací může ovlivnit tento progess.
Nanášející smrtelný, mutace v endolethin-B receptoru: Vymazání v genu pro endolethin-B receptor, (mutace nazvaná volala nanášející smrtelný (sl )) vede k problémům v melanocyte a eneterickém buněčném rozptýlení v krysách. Toto vede k depigmentation čelového (požáru) a nedostatek nervového spojení k dvojtečce, které prostředky bowel nemůže být odstraňován. Neschopnost odstraňovat bowel je podmínka volala megakolon nebo megacecum, a to je fatální (Dembowski et al. 2000, Tsaur et al. 1997, Kunieda et al. 1996, Gariepy 1996, Vyhrál et al. 2002).
- Sl = normální
- sl = nanášející smrtelný
Poznámka: myši mají srovnatelnou mutaci, která ovlivňuje Endolethin B, nazvaný strakaté nanášení (s). Oni také mají oddělenou mutaci, která omráčí Endolethin 3, s podobnými účinky. Matouce, toto Endolethin 3 mutace v myších je nazvaná smrtelné nanášení (ls ), který je různý od nanášející smrtelný (sl) v kryse. Whoever jmenoval tyto mutační potřeby mít jeho hlavu prozkoumanou.
Pro více na endolethin-B receptoru a megakolonu v myších, vidět Zhua et al. 2004, a pro více na myšových modelech Waardenburg syndromu, vidět Tachibana et al. 2003.
Bílé nanášení, mutace v Kit bílkovině: Oddělená mutace, nazvaný bílé nanášení (Ws) omráčí Kit bílkovina, tyrosine kinase transmembrane receptor, který je vytvořený c-kit genem. Mutace je 12-base základu vymazání v c-kit genu (Tsujimura et al. 1991 ). Výstrojová bílkovina má širokou rozmanitost funkcí! Kit je zapojený do vývoje krevních kmenových buňek (předchůdcové červených a bílých krvinek ), melanoblasts a pravěká bakterie buňky a melanoblast stěhování (Horie et al. 1991 ). Tak plení výstroj bílkovina bude mít paletu efektů, zahrnující: depigmentation jistých oblastí (zvýší k a zahrnující úplně bílý s černými očima ), a někdy chudokrevnost, nedostatek žírných buňek (a proto nedostatky v histaminu a serotonin ), reprodukční problémy a hluchota (Kitamura et al. 1994, Hoshino 2000, Sugimoto 1995 ).
Tito nejsou jediné role Kit bílkoviny, nicméně. Kit je také používán v střevních zkracováních: Zvláštní buňky v střevní zdi, nazvané intersticial buňky od Cajal (ICC) tvořit elektrické pomalé vlny v gastrointestinální ploše. Tyto pomalé vlny regulují frekvenci střevních svalových zkracování, tak ICC je kritický pro normální motility malého střeva. ICC syntetizuje Kita, produkt c-kit. Krysy s bílou, která všímá si mutace neprodukují Výstroj v jejich střevech, a neukazují elektrické pomalé vlny. Toto skončí abnormálním zkracováním a megakolonem (Takeda et al. 2001 ).
- Ws = bílé nanášení
- ws = normální
Poznámka: Bílé nanášení (Ws) v kryse je podobný, ale ne totožný, k bílé nanášení (W) v myši. Oba jsou mutace v c-kit genu.
Příklady: megakolon je známý být příležitostně spojený s plápoláním v krysách. Někteří plápolali linky, takový jak zastřený (depigmented čela a strany, někdy zvláštní prohlížel si) může mít vyšší dopad megakolonu.
Poznámka, nicméně, ten megakolon může být způsobený jinými faktory také (e.g. oddělený druh vrozeného megakolonu neznámé příčiny, Lipman et al. 1998 ), že ne všechny požáry jsou způsobené u všiml si smrtelného nebo bílého nanášení geny, a že ne všichni plápolali / depigminted krysy mají megakolon!
Lidé a jiný druh: Lidé mají mutace v barvivovém buněčném stěhování příliš, volal Waardenburg syndrom. Tam je mnoho typů Waardenburg syndromu, každý způsobil mutací v různém genu (PAX3, MITF, EDNRB, EDN3 a SOX10 ). Nejhroznější forma Waardenburg syndromu, volal Waardenburg Typ 4 nebo Waardenbug-Shah, zahrnuje depigmentation, oční barva anomálie a megakolon megakolonu (je nazvaný Hirschprung-ova nemoc u lidí ).
Endolethin B: V koních, mutace v Endolethin B vede k Smrtelnému Bílému Hříběti Syndrom (LWFS) (Santschi et al. 1999 ). Pro více, vidět Endolethin Receptor stránkovat v Online Dědičnosti v Mužské databázi.
Kit gen je zapletený do bílé barvy některých psích chovů, kde to je nazvané nanášení (s). Nanášení má několik alleles v psech. Dominantní forma (S) produkuje pevný kabát, někdy s trochu bílou kolem prstů, hruď a břicho. Irské nanášení s (i) allele produkuje bílá známkování na foreface, krk, nižší údy, hruď a břicho (e.g. Boston Terrier a Basenji ). V těchto zvířatech, barvivové buňky nikdy dělají to kolonizování míst na krku (Cattanach 1999 ).
Strakatý nanášející gen s (p) produkuje širší distribuci white (Cocker křepeláci, Ukazatelé ). Nejextrémnější allele, s (w) je najitý v Dalmatians, Angličtina Setters, bílý Býk Terriers, a bílí Boxeři, kde to produkuje štukovou omítku, někdy s zabarvenými body. V těchto zvířatech, většina kabátu depigemented, ale několik barvivové buňky mohou dělat to kolonizujícím místům (pozoruhodně kolem očí a uší ), který množit se a produkovat tmavé skvrny (Cattanach 1999 ).
Tento extrém s (w) allele je zodpovědný za vysoký výskyt hluchoty v Dalmatians (20-30 % od Dalmatians být sluch poškozený --bilaterally nebo jednostranně hluchý ). Poznámka, nicméně, to Dalmatians s barevnými místy na jejich uších mít nižší dopad hluchoty, který ukáže, že jestliže barvivové buňky dělají to uchu sluch inklinuje k být normální (bohužel, místa jsou zvažována chyba v Dalmatians, tak chovatelský chov proti místům, a od této doby být bezděčně vybírající pro hluchotu -- bummer pro psy ). Interestingly, Boston Terriers je povolený mít hlavu místa, a oni mají nižší dopad hluchoty (Cattanach 1999 ).
Mutace v Kit genu je také zodpovědný za barevný vzor Hereford dobytku, ale není spojován s hluchotou. Poznámka, nicméně, ten Hereford dobytek všichni mají zabarvené uši.
Mutace v melanosome formaci
Problém v počtu řádků buňek, které vedou k melanosomes: red prohlížela si roztok allele
Melanosomes je malý malý vesicles najitý vnitřek barvivová buňka. Barviva jsou shromážděná v těchto malých melanosomes, který být pak transportovaný k okraji barvivové buňky a sázet jejich barvivo v rostoucích vlasech.
Melanosomes je vlastně část rodina příbuzných " buněčných orgánů " (organelles) (Orlow 1995, 1998 ), to zahrnuje lysosomes a krevní destička hustá granules. Lysosomes málo vesicles vnitřní buňky, které obsahují enzymes zapojený do rozbíjejícího se dole metabolites (odpadu ). Krevní Destička husté granules jsou najité v krevních krevních (destičkách, které oni uloží a vylučují adenosine nucleotides a serotonin ). Defekty v krevní destičce husté granules vedení k chudé krvi, která se sráží a prodloužily krvácení.
Tyto tři druhy organelles, melanosomes, lysosomes a krevní destička hustá granules, všichni sestupují od obyčejného předcházejícího organelle. Proto, nějaká mutace, která ovlivní tohoto obyčejného předka ovlivní potomky. Recesivní red-eyed prohlížela si roztok mutace (r) má jen tento účinek.
Červená prohlížela si roztokovou mutaci překáží normálním vývojem těchto organelles. Toto vede k abnormální dopravě melanosomes uvnitř barvivové buňky, které příčiny redukovaly barvivový deposit v vlasech a prohlížely si -- od této doby červené oči a bledá kožešina. Homozygotní krysy s red-eyed prohlížela si roztok také mít abnormální destičkovou funkci, nazýval Krevní Destičku Ukládacím Kalužovým Nedostatkem (SPD) (LaVail 1981, Prieur 1984 ). V krysách s SPD, krevní destičky mají vadné vylučování srážejících se prostředníků, který vede k hojnému vypouštění (Raymonda a Dodds 1975, Tschopp a Baumgartner 1977, Kirchmeier et al. 1990, Magro et al. 1992).
Red-eyed roztok je docela různý od pink-eyed prohlížela si roztok, thoug zvířata mohou mít podobný vzhled (ačkoli rr krysy mají rudé hnědé oči, chvíle pp krysy mají opravdově růžové oči (LaVail 1981 )). Geneticky, nicméně, tito jsou docela zřetelné mutace, které mají velmi různé účinky.
Příklady: Jinak agouti krysa homozygotní pro red-eyed prohlížela si roztok bude zlatý opalovat se nazvaný kolouch (Prieur 1984)
Poznámka: Červená prohlížela si roztok agouti není jediný způsob, jak sehnat kolouchu barevnou krysu. Tam je také oddělit se kolouch mutace (f ), který redukuje zabarvení v jak černých tak modrých zvířatech, ačkoli jeho buněčný mechanismus je neznámo. Kolouch na černé kryse produkuje kávu hnědé zvíře, chvíle kolouch na modrém zvířeti produkuje koloucha zvířecí (Hrad a Král 1947 ).
Jiné účinky: Kolouch (rr na agouti) zahalené krysy jsou použité značně v výzkumu, a mít celý seznam sdružených nepořádků:
- hypertension (Rudofsky a Magro 1982, Kuijpers et al. 1986, Kuijpers a Jong 1986) vede k:
- proteinuria (bílkovina v urine) (Kuijpers et al. 1986, Kuijpers a Jong 1986)
- ohnisková glomerulární skleróza (scarring ledvinové tkáně) (Kreisberg a Karnovsky 1978)
- ledvinová porucha (de Keijzer et al. 1989)
- změněná citlivost serotonergic mechanismů v centrálním nervovém systému (e.g. Gudelsky et al. 1985, Wang et al. 1988 ). Pro více detailů, vidět Hulilhan-Giglin, 1992, 1993, Chen a Lawrence 2000.
- alkoholismus (Daoust et al. 1991, Overstreet et al. 1992, Rezvani et al. 2002).
- vysoko sociální úzkost a nízká agrese (Kantor et al. 2000).
Fawn zahalené krysy byly použité jako zvířecí model pro lidské psychiatrické nepořádky, které zapojují anomálie do serotonin funkce, takový jako:
- deprese (Overstreet et al. 1992, Rezvani et al. 2002)
- úzkost (Altemus 1994)
- posedlý poutavý nepořádek
- jedící nepořádky
Jen destičkový ukládací nepořádek diskutoval o nahoře, a serotonin uptake nepořádek (Tobach et al. 1984) byli ukázaní být způsobený červenou prohlížel si roztokový gen (Hamada 1997, Fugimori et al. 1998, Prieur 1984 ). Mnoho tyto jiné nepořádky mohou být způsobené jinými geny, který přišli k být spojován s těmito laboratorními počty řádků krys (koloucha Overstreet a Rezvani 1996, Overstreet et al. 1999, Rezvani et al. 2002).
Lidské analogie: Tam být přinejmenším 15 myši analogie, zahrnující: lehké ucho,maroon, bledý, perlový a rubínový prohlížel si. Jako když red-eyed prohlížela si roztok, tyto podobné mutace ovlivní melanosomes, destičkové uskladnění granules a lysosomes. Nicméně, žádná z těchto podobných mutací být přesně stejná mutace jako red-eyed prohlížela si roztok. Jinými Slovy, tyto analogie ovlivní stejný proces ale v různých cestách (Nguyen et al. 2002, Prieur 1984 ).
Tam být několik lidské analogie, které ukazují destičkový ukládací nedostatek a depigmentation:
Hermansky-Pudlak syndrom (HPS ). Jednotlivci s HPS mají rozsah depigmentation, od bílých vlasů a kůže k hnědým vlasům a kůži kvůli mnoho pihám. Jednotlivci s HPS snížili ostrost vidění a problémy (plíce plicní fibróza) a střevní problémy (granulomatózní kolitida ). HPS je způsobený problémem v bláně bílkoviny tří organelles zmínily se, že nahoře (melanosomes, lysosomes a destičkové uskladnění granules) který skončit vadnou dopravou. Tam jsou přinejmenším tři různé druhy HPS (Huizing a Gahl 2002 ). To je vzácné v nejvíce lidských populacích ale je nejvíce obyčejný druh albinism v Portoriku. Myš homologue od HPS je bledé ucho.
Chediak-Higashi syndrom (CHS ). Lidé s CHS ukazují hluboký depigmentation kůže, hrozné recidivující nákazy, množení lymfatické tkáně (, která lymphoproliferative nepořádek) a numbness extremeties (postupného periferního neuropathy ). Jednotlivci s CHS mají obra melanosomes v jejich melanocytes a obru lysosomes v jejich bílých krvinkách (leucocytes ). Toto je vážný stav a mnoho lidé s CHS umřou předčasně. CHS je také najit v norku, dobytek, myši a kočky. Myš homologue od CHS je beige. Tam je také přinejmenším jeden příklad v literatuře z beige krysy s CHS symptomy (Ozaki et al. 1998).
Mutace v barvivové dopravě a depositition
I. Přepíná Mezi lehký a tmavý na jediných vlasech: černý a agouti gen
Melanocytes produkuje dva druhy barviva, osmahnout černý eumelanins a červený žlutý phaeomelanins. Poměrné podíly těchto barviv jsou regulované u:
(1) alpha-MSH, který sváže k melanocortin receptor volal MCR1 na melanocytes a povzbudí je produkovat hnědý černý eumelanins povzbuzováním výroby tyrosinase, a
(2) Agouti bílkovina, který zabrání MSH od vázání k MCR1 a skončí výrobou červeného žlutého phaeomelanins (Lu et al. 1994).
Agouti má oddělenou cestu ke akci také: Agouti zabrání melanogenesis a obecně redukuje syntézu obou barviv (Graham et al. 1997).
- A = aguoti
- = non-agouti
Tak, krysy s agouti bílkovinou střídají se mezi hnědou černou a červenou žlutou barvivovou výrobou, produkování odstupňovaných vlasů, který je divoký typ phenotype Norské krysy. Krysy bez agouti bílkoviny nestřídají se. Oni produkují eumelanins nepřetržitě a jejich vlasy jsou stejná temná barva délka šachty.
Příklady: Krysy s aa mít pevné vlasy (s žádnými jinými modifikátory, tito jsou black krysy ), krysy s A odstupňovali vlasy, nazvaný agouti.
Jiné efekty: Agouti bílkovina také předejde MSH od vázání k jeho melanocortin receptorům na nervových buňkách (Lu et al. 1994, Willard et al. 1995 ). Tyto melanocortins jsou silné neuromodulators, které mají různorodé účinky na savce chování a fyziologie, takže non-agouti krysy jsou klidnější a snadnější ovládat se než agouti krysy (Keeler 1942, Cottle a Cena 1987 ). Behavioral rozdíly mezi agouti a nonagouti krysami jsou pravděpodobně kvůli pravidlu od MSH a jeho účinky na mozek a následující chování.
Agouti gen proto má dopad na chování a může byli důležití v domestikace krysy. Osmdesát procento domácí laboratoře krysí napětí jsou homozygotní pro nonagouti allele (vidí Cenu 2002 p. 16-17 pro diskuzi ).
Další vedlejší účinek vyskytuje se v mutantu s přes výraz agouti v myších. Agouti sváže k nervovým melanocortin-4 receptorům (MC4) v mozku, který být zapojený do příjmu potravy a homeostasis (Skuladottir 1999 ). Myši s přes výraz Agouti být žlutý (od exkluzivní výroby phaeomelanins) a otylý od zvýšeného příjmu potravy.
Lidské analogie: Lidé mají agouti analogie nazvaná agouti signálovou bílkovinu, ASIP, ale jeho role je uboze definovaná. To je vyjádřené nejvíce vadipose (tuk) tkáň kde to může znepřátelit si jeden z melnocortin receptorů. To může hrát roli v energii homeostasis a a možná lidské zabarvení (Voisey & Daal 2002 ).
Zajímavější látka okolo MCR1-MSH ... jak vy byste mohli čekat, mutace v MCR1 receptoru by měla zajímavé důsledky pro zabarvení. MCR1 sváže MSH, který způsobí výrobu tmavého eumelanin. S non-functional MCR1, MSH nemůže svázat, a melanosomes produkuje jediné světlo, žlutý nebo červený phaeomelanins. Mutace v rozšíření, gen ten kódy pro MCR1, nejsou najití v krysách. Ale oni jsou najití v množstvích jiného druhu, takový jako myši, který produkuje žlutý kabát vybarvené (myši, rozšíření je nazvaný yellow (e)) (Robbins et al. 1993). Rozšíření mutace v koni produkují kaštan obarvit (Marklund et al. 1996). Rozšíření je také najit v červených liškách, a v psech takových jako žlutý labradors, zlatý retrievers a Irský setters (Newton et al 2000; Everts et al. 2000).
Jiné mutace v MCR1 přimějí to, aby byl hyperaktivní, vázání MSH neustále a vede k plynulé výrobě tmavého eumelanin. Černá černého panthers je způsobená takovou mutací (Robbins et al. 1993 ), jako ně černá černé ovce (Vage et al. 1999 ). Hyperaktivní MSH mutace jsou najité v myších příliš, v formě z somber a tabák tmavnoucí alleles (Robbinse et al. 1993).
MCR1 mění se sezónně v nějakém druhu, který je co způsobí přechod od bledých zimních kabátů k tmavému jaru a letním kabátům v některých zvířatech.
U Lidí, mutace v MCR1 receptorech přimějí melanocytes, aby syntetizoval červený žlutý phaeomelanin, který produkuje rudé vlasy. Tam být 6 známé mutace v MCR1 který měnit se v jak hodně oni omráčí MCR1-OVU funkci. Rusovlasí jednotlivci jsou obvykle homozygotes nebo složený heterozygoes (, který posessing dva různé druhy MCR1 mutace ). Dva alleles pro non-functional MCR1 bude produkovat jasné rudé vlasy. Lidé s jen jeden měnil MCR1 allele nebo alleles, které jen částečně poškodí MCR1-OVU funkci, může ukazovat měnící se odstíny červené nebo žádné červené vůbec. Asi 50 % bílých lidí nést měnil MCR1, který je myšlenka být významně spojený s světlou kůží dokonce jestliže to neprodukuje rudé vlasy (Ha a Rees 2001, Rees 2000, Schaffer a Bologna 2001 ).
Jak to dopadá, phaeomelanins nemají jak dobrý v chránění kůže od ultrafialových paprsků jako eumelanins být, který je proč rusovlasí lidé jsou více náchylní k rakovině kůže.
- Pro více na rudých vlasech: Red Hair Genetika
II. Less barvivo vsazené v vlasech skončí lehčím kabátem: ředit gen
Jakmile barviva jsou vyrobená, oni musí být transportovaní k vlasům. Barvivové částečky, volaný granules, být vysyntetizovaný v málo vesicles nazvaném melanosomes, který být transportovaný přes dendrites melanocyte k komínu rostoucích vlasů. Doprava je vlastně opravdu čistý ... melanosomes jsou nesené málo molekulárními nohami (myosin 5, actin závislá motorová bílkovina) podél malého odvětvování molekulárních " žebříků " (actin vlákna) obsáhnutý v projekcích mimo melanocyte (dendrites) k buněčnému okraji. Jestliže tento přepravní systém je zasažený, doprava k buněčnému okraji není normální.
Krysy s zředěnou mutací (volaly Myo5a (d) v literatuře) produkce normální typy a množství barviva v jejich melanocytes ale dopravě těchto barviv k vlasové šachtě jsou přerušená kvůli mutaci v myosin 5. Mnoho melanosomes s jejich barvivy jsou vězel v buněčném centru, neschopný být přenesený ven k buněčnému okraji. Tak méně barvivo je včleněné v vlasech, a když to je včleněné to inklinuje k být vsazený v shlukách. Toto dává kabát bledá, zředěná barva (Wu et al. 1998, Wei et al. 1997).
- d (také volal MyoVa, Myo5a) = ředit
- D = normální
Tam je několik alleles zředěného genu, takový jak zředěný smrtelný a zředěný opisthotonus (dop ), který skončit hroznou neurologickou defektní (ataxií, záchvaty) a někdy smrt v homozygotním státu (Futaki et al. 2000; Ohno et al., 1996; Dekker-Ohno 1993 ).
Příklady: Krysy, které jsou geneticky černé ale mají zředěnou mutaci jsou břidlice šedá nazvaný modrý.
Poznámka, nicméně, že tam být mnoho různé mutace, které dají zvednou se k modré krysy v oblíbené kryse přepychové, tak to není jasné která oblíbená krysí mutace (jestliže některý) odpovídá známé mutaci v MyoVa v laboratorních krysách. Také poznamenat, že názvy různých odstínů blue v mazlíčku krysy a Mendelian zápisy určily je mezi krysí chovatele může lišit se mezi oblíbenou krysou organizace a země.
Jiné účinky: Barvivová doprava není jediná postižená věc když myosin 5 je poškozený. Myosin 5 je také používán v vypracování a udržování buněčných procesů melanocytes: homozygotní d krysy melanocytes s méně a tenčí dendritické procesy. Myosin 5 je také zodpovědný za táhnutí endoplazmatického reticulum (ER) k buněčnému okraji Purkinje buňek (velkých, důležitý neurons v mozku) kde to produkuje toky vápníku. Jestliže myosin 5 je přerušený, to nemůže táhnout ERA k buněčnému okraji, toky vápníku jsou přerušené, který srazí se Purkinje buňka je vzrušivost, který skončí neurologickými deficity jak viděný v část hroznější zředěné mutace (vidí Tab et al. 1998; Takagishi 1998; Hurvitz et al. 1993 ).
Lidské analogie: Lidský protipól zředěné mutace je najitý v Griscelli Prunieras syndromu, unikátní autosomální recesivní nemoc (Westbroek et al 2001 ). Tam jsou dva typy. Jednotlivci s Griscelli typem 1 mít mutaci v myosin 5 a být charakterizován částečným albinism, stříbrné plavé vlasy odbarvení a neurologické deficity. Jednotlivci s Griscelli typem 2 mít mutaci v další dopravní bílkovině, volal Rab27a, a oni mají příznaky typu 1 nebo více primární volby immunodeficiency (Kleina et al. 1994 ). Melanocytes lidí s Griscelli symdromes mít krátký, podsaditý dendrites a obsahovat zralý melanosomes, ukazování převodového bloku k obklopujícímu keratinocytes.
- Více na albinism u lidí
- Griscelli syndrom: http:// www.ncbi.nlm.nih.gov / entrez / dispomim.cgi?id =214450
- Dobrá strana na myosin 5: http:// www.bms.ed.ac.uk / výzkum / jiní / smaciver / Myosin %20V.htm
Mutace v barvivové syntéze
I. Hasák pravý u začátku: Chinchilla mutace
Jedna z nejčasnějších mutací v barvivové výrobní stezce je mutace v enzyme tyrosinase, který přestaví barvivového předchůdce tyrosine do příštího kroku. Gen pro tyrosinase je pojmenovaný chinchilla. Jestliže mutace v chinchilla genu skončí kompletně nonfunctional tyrosinase enzyme pak zvíře bude neschopné produkce nějaké barvivo kdekoli v tělu. Zvířata s touto mutací jsou nazvaná albíni.
Chinchilla gen má jiné známé mutace, nicméně, a mnoho je skončit polořadovkou funkční tyrosinase. Tito produkují zvířata s zředěnou barvou srovnanou k těm s normálním tyrosinase. V acromelanic verzi, polořadovka funkční tyrosinase je velmi křehká a teplota závislá. Zvýšit Teplotu příliš mnoho a tyrosinase zlomí se. Krysy a jiná zvířata s teplotou citlivou tyrosinase jen produkují barvivo v chladnějších oblastech jejich těl: extremeties takový jako nos, uši, nohy a ocas. Zvířata s těmi teplota citlivé mutace jsou siamese nebo himalayan. Pro specifický molekulární popis z jak tyrosinase vytvořený achromelanic mutací liší se od normálního tyrosinase, vidět Kwon et al. 1989.
Tady být část mutace v chinchilla genu pro tyrosinase:
- C = plná barva
- C (ch) = Chinchilla
- c (h) = Acromelanism (siamese)
- c = Albín
Příklady: C (ch) C (ch) produkuje chinchilla, ale toto je (pokud já vím to) unheard z v krysách. Krysy s cc být albín, a c (h) c (h) krysy jsou známé jak siamese. Kombinace c (h) c produkuje zabarvení střední mezi albínem a siamese a je známý jak himalayan.
Jiné efekty: nedostatek melaninu barvivo v oku a zrakových nervech vede k abnormálnímu vývoji a funkci, a od této doby chudá vize a někdy slepota (Balkema a Drager 1991 ). Nepřítomnost fovea, nystagmus, strabismus a redukovaná ostrost vidění je obyčejná v všech druhách albinism. Pro více na jak bílé krysy poznají svět, vidět článek opravňovaný "co krysy vidí?"
Lidské analogie:albinism vyskytuje se v lidech, příliš, kde to je nazvaný oculocutaneous albinism typ 1 (OCA1 ). Tam být asi 60 mutace, které ovlivní tyrosinase gen u lidí (Oetting & Král 1993, Spritz 1994 ). Jak hodně barvivoví jednotlivci produkce závisí na jak impared tyrosinase enzyme je -- barviva výraz může toulat se od úplně chybějící (bílé kůže, bílé vlasy, a depigmented oči) k téměř normální.
Tam jsou také lidské obdoby himalayan nebo siamese mutace, varianta oculocutaneous albinism typu 1 (OCA1 ). Rádi siamese zvířata, lidé s teplotou citlivou tyrosinase produkují bílé vlasy v teplých oblastech jejich těl a tmavějších vlasů v chladnějších oblastech. Oni mají bílé axilární (podpaží) vlasy, téměř bílé skalpové vlasy, lehká hnědá paže vlasy a tmavá hnědá noha vlasy a žádná celková kůže barvivový (Král et al. 1991; Giebel et al. 1991; Oetting & Král, klinická škála albinism u lidí,).
- Více na oculocutaneous albinism u lidí
- Také vidět Boissy 1997.
II. Chudé buněčné prostředí pro barvivovou syntézu: pink-eyed prohlížela si roztok mutace
Barvivová syntéza vyskytuje se v melanosomes: Tyrosine vstoupí do melanosome a enzyme tyrosinase, který catalyzes to do dopaquinone. Později, srážet dolů eumelanin stezku, tyrosinase-related-proteins změní hnědé barvivo na černý eumelanins. Tyto chemické reakce mohou být povznesené nebo poškozené změnami v vnitřním pH melanosome.
The p místo kódy pro bílkovinu umístily na eumelanosome's bláně, hradlová (transportérová bílkovina) to zasvětí molekuly do buňky. Brána vypadá, že pomůže regulovat pH melanosome tím, že vpustí anions (různá brána připustí H + ). Mutace v p místo ovlivní tuto bránu, měnění normálního, kyselého pH k neutrálnějšímu prostředí. As p bílkoviny nejsou najité na phaeomelanosomes, phaeomelanin výroba není zasažená mutacemi v p.
Kyselé prostředí v melanosome je požadované pro normální tyrosinase aktivitu (Oslnivou et al. 2001. Strum 2001. Puri et al. 2000 ). Když melanosomes být kyselý, oni produkují více barviva, s preferenčním zvyšováním tmavého eumelanins. Když oni jsou neutrální, oni produkují méně eumelanin ale phaeomelanin je relativně nedotčený (Oslnivý 2001 ). Tam je také nějaká otázka z p gen, který je tyrosine transportér (Rinchik 1993 ), ale důkaz je smíšený.
Tak p allele zředí kteroukoliv barvu to je najité s, tím, že redukuje černá hnědá barviva a odchází červená žloutne sám. Čistý výsledek, v krysách, je redukce v tmavých eumelanin barvivech, končí bledým kabátem a růžovými očima.
- P = normální zabarvení
- p = pink-eyed prohlížela si ředit
Příklady: v agouti kryse, karafiát prohlížel si roztok zředí tmavou skupinu na každých vlasech k bledé barvě, končí celkovou kombinací někdy volal jantar. Rovná černá je zředěná k bledě yellow který je někdy volán champagne.
Jiné efekty: málo, abnormálně zformovaný melanosomes.
Lidská analogie: Normální variace v lidském p místě způsobí normální kůži barevnou variaci mezi různé etnické skupiny.
Silný efekt krysy je růžový oční roztok p mutace má jeho člověka analogového v oculocutaneous albinism typu 2, (OCA2 ). OCA 2 je nejvíce obyčejný druh albinism u lidí, obzvláště mezi lidi Afrického svahu. Characeristics je velmi proměnlivý, nicméně, vytyčování od minimální mírnit zabarvení vlasů, kůže a duhovka. Barvivo může být lokalizované v pihách. Jednotlivci Afrického svahu s OCA2 inklinují k mít žluté vlasy, bílá kůže s lokalizovaným barvivem oblasti a duhovky, které jsou částečně nebo kompletně zabarvený s opalovat melanin. The p gen u lidí je lokalizovaný na segmentu chromozómu 15, který je vymazaný v lidech s Prader-Willi a Angelman syndromy, který může vysvětlit, proč jednotlivci s těmito syndromy hypopigmented vlasy, oči a kůže (Rinchik et al. 1993, Oslnivý et al. 1994).
III. Poslední krok v barvivové výrobě, která nestane se: brown mutace
Poté, co dopaquinone, barvivová stezka rozdělí se do dva. Jedna větev vede k eumelanin a jiným vedením k phaeomelanin. Jedna z mutací to ovlivní výrobu eumelanin je mutace v brown gen, které kódy pro enzyme TYRP1, který catalyzes konečný krok eumelanin syntézy: přestavování hnědého barviva k black. S normální TYRP1, konečný krok vezme místo a zvířata produkují černý eumelanin. Mutace v TYRP1 prostředkách tento konečný krok nemůže stát se, a zvířata s touto mutací produkují hnědý eumelanin. Tam jsou přinejmenším dva druhy TYRP1 mutací: jeden je genová mutace v TYRP1 který poskytne tomu non-functional, jiný je mutace jinde to způsobí TYRP1 být vytvořený v nízkých kvantitách.
- B = black
- b = zhnědnout (také věděl to jako TYRP1 (b ))
Příklady: Krysy homozygotní pro hnědou mutaci jsou nazvané čokoláda, v psech toto je často voláno játra. Agouti krysy homozygotní pro brown mají brown místo black eumelanin pruhy na jejich vlasech a být někdy nazvaný čokoláda agouti.
Jiné účinky: TYRP1 mutace také ovlivní buněčné rozšíření a melanosomal zrání (Sarangarajan et al. 2000).
Lidské analogie: mutace v TYRP1 jsou najité u lidí, příliš, kde oni produkuje oculocutaneous albinism typ 3 (OCA3 ). U Lidí jak v myších a krysách, TYRP1 catalyzes eumelanin syntézu, ale u lidí to může mít další role, takový jako udržování stability tyrosinase a upravování jeho aktivity. To také maintainins melanosome strukturu a ovlivní melanocyte rozšíření a buněčnou smrt (Sarangarajan & Boissy 2001, Sarangarajan et al. 2000).
Oculocutaneous albinism Píší 3, také věděl to jak rufous oculocutaneous albinism, xanthism nebo xanthous albimisn, je obyčejný autosomální nepořádek najitý v černoších (Manga 1997 ). Lidé s rufous albinism produkce normální phaeomelanins (červené a žloutne) ale neprodukují černý eumelanins, jen ty hnědé. První popis osoby s xanthous albinism byl African-American dvojčete chlapec, který měl lehkou hnědou kůži, lehké hnědé vlasy a modré / šedé duhovky zatímco jeho bratrské dvojče mělo normální zabarvení. Více jednotlivců bylo popisováno od té doby pak, takoví jednotlivci mají červenou k rudému hnědému zbarvení kůže a vlasů a rudý hnědý, mírně průsvitné duhovky; photophobia a nystagmus je slabé pivo a ostrost vidění je normální nebo téměř normální.
- Více na oculocutaneous albinism u lidí
Online zdroje:
- Albinism Databáze: http:// www.cbc.umn.edu / tad /
- Albínské geny: http:// www.cbc.umn.edu / tad / genes.htm
- Fakty o Albinism: http:// www.cbc.umn.edu / iac / newfacts.htm
- oculocutaneous albinism u lidí http:// www.cbc.umn.edu / iac / mmm.htm
- Myš, která pigmentary mutovanou úschovnu: http:// www.sghms.ac.uk / depts / anatomie / strany / MRHOLDINGS.HTM
- Klinická škála albinism u lidí. Univerzita od Minnesoty. http:// www.cbc.umn.edu / iac / mmm.htm
- Nervové Hřebenové Zabarvení Mutanti: http:// vtpb-www.cvm.tamu.edu / Larr _Myš / pigment.html
Více mutací přijít jako já zkoumám je!
